El proceso por el cual las células degradan las moléculas de alimento para obtener energía recibe el nombre de respiración celular.
La respiración celular es una reacción exergónica, donde parte de la energía contenida en las moléculas de alimento es utilizada por la célula para sintetizar ATP. Decimos parte de la energía porque no toda es utilizada, sino que una parte se pierde.
Aproximadamente el 40% de la energía libre emitida por la oxidación de la glucosa se conserva en forma de ATP. Cerca del 75% de la energía de la nafta se pierde como calor de un auto; solo el 25% se convierte en formas útiles de energía. La célula es mucho más eficiente.
La respiración celular es una combustión biológica y puede compararse con la combustión de carbón, bencina, leña. En ambos casos moléculas ricas en energía son degradadas a moléculas más sencillas con la consiguiente liberación de energía.
Tanto la respiración como la combustión son reacciones exergónicas.
Sin embargo existen importantes diferencias entre ambos procesos. En primer lugar la combustión es un fenómeno incontrolado en el que todos los enlaces químicos se rompen al mismo tiempo y liberan la energía en forma súbita; por el contrarío la respiración es la degradación del alimento con la liberación paulatina de energía. Este control está ejercido por enzimas específicas.
En segundo lugar la combustión produce calor y algo de luz. Este proceso transforma energía química en calórica y luminosa. En cambio la energía liberada durante la respiración es utilizada fundamentalmente para la formación de nuevos enlaces químicos (ATP).
La respiración celular puede ser considerada como una serie de reacciones de óxido-reducción en las cuales las moléculas combustibles son paulatinamente oxidadas y degradadas liberando energía. Los protones perdidos por el alimento son captados por coenzímas.
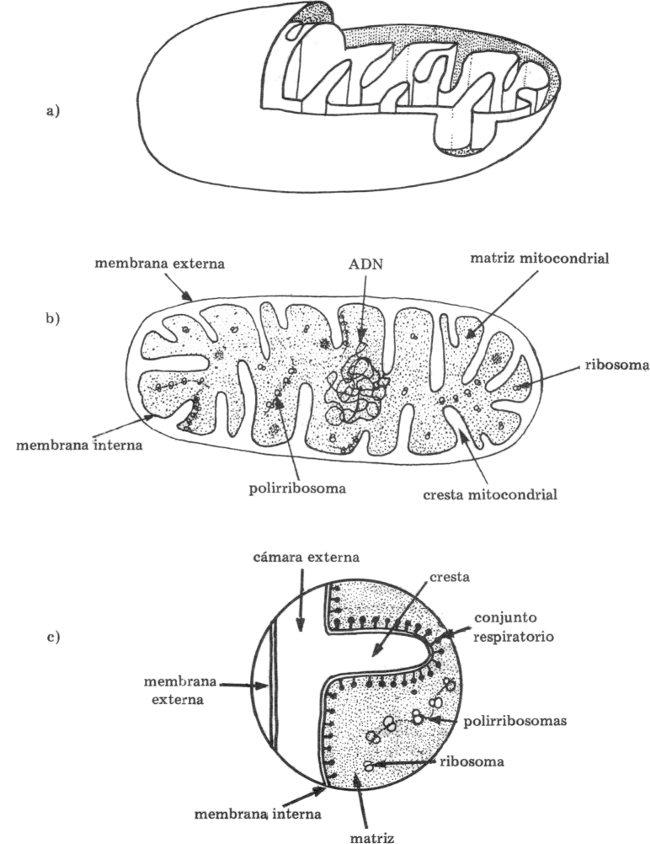
La respiración ocurre en distintas estructuras celulares. La primera de ellas es la glucólisis que ocurre en el citoplasma. La segunda etapa dependerá de la presencia o ausencia de O2 en el medio, determinando en el primer caso la respiración aeróbica (ocurre en las mitocondrias), y en el segundo caso la respiración anaeróbica o fermentación (ocurre en el citoplasma).

A continuación les dejaremos una animación de la respiración celular: